
Effects of Qigong on the Nervous System
(9a)
by Martin Eisen, Ph.D.
Introduction
Qigong produces physiological effects on the nervous system. To appreciate and understand these effects, some background material on the nervous system is required and will be presented below.
More details can be found in the references. Qigong’s effects on the nervous system will be discussed in subsequent articles.
2. Cells of the Nervous System
The nervous system is composed mainly of nerve cells or neurons and glial cells or neurogllia or glia. Neurons propagate electrical signals by electrochemical means of ion transfer across the cell’s membrane. There are about 100 billion neurons in the brain. Glia are more numerous than neurons.
Glial Cells
About 90 percent of the brain’s cells are glial (meaning “glue”) cells, which don’t carry nerve impulses.
The four main functions of glia are to surround neurons and hold them in place, to supply nutrients and oxygen neurons, to insulate one neuron from another by manufacturing myelin, to destroy pathogens and remove dead neurons. They also modulate neurotransmitters. Some types of glial cells are Schwann’s Cells (produce myelin), Satellite Cells, Microglia,Oligodendroglia, and Astroglia. Neuroglia guide neurons during fetal development.

Figure 1. Neuron with a myelin sheath
The word “neuron” was coined by Heinrich Wilhelm Gottfried von Waldeyer-Hartz in 1891. Neurons vary in size from 4 microns (.004 mm) to 100 microns (.1 mm) in diameter. Their length varies from a fraction of an inch to several feet. The neuron (see Fig. 1) consists of a cell body (or soma) with branching dendrites (signal receivers) and a long projection called an axon, which conduct the nerve signal away from the soma. The cell body contains the neuron’s nucleus (with DNA and typical nuclear organelles). Dendrites branch from the cell body and receive messages. Myelin coats and insulates some axons (except for periodic breaks called nodes of Ranvier), to increase the transmission speed along these axons. Myelin is manufactured by Schwann’s cells, and consists of 70-80% lipids (fat) and 20-30% protein. Bundles of axons are known as nerves. A typical neuron communicates with 1,000-10,000 other neurons, muscle cells, glands, etc.
Neurons cannot regrow after damage, except for neurons from the hippocampus). Fortunately, there are about 100 billion neurons in the brain and they can form new connections.
Types of Neurons
Different types of neurons are found in different parts of the body (see Table 1). They all propagate nerve signals, but differ in the number of processes emanating from the cell body.
| Type | Function | No. of Processes | % of all Neurons | Example |
| Sensory or Bipolar | Carry messages from the body’s sense receptors (eyes, ears, etc.) to the CNS. | 2 | .9 | Retinal cells, olfactory epithelium cells |
| Motoneurons or Multipolar | Carry signals from the CNS to the muscles and glands. | Many | 9 | Sinal motor neurons, pyramidal neurons, Purkinje cells |
| Interneurons or Pseudopolar (Spelling) | Form all the neural wiring within the central nervous system | 2 axons. Go to spinal cord & skin or muscle. | 80.1 | Dorsal root ganglia cells |
Table 1. Neuron Types
Synapses
The message or nerve impulse relayed from one neuron to another neuron or cell is by means of the action potential carried by the neuron’s axon.The electrical impulse arrives at the axon terminal.There is commonly a very narrow cleft (about 20 nm in width) between the neurons or the neuron and cell called a (chemical) synapse.Most synapses connect axons to dendrites, but there are also other types of connections, including axon-to-cell-body, axon-to-axon, and dendrite-to-dendrite.The presynaptic neuron has small membrane containers, called vesicles, containing chemical compounds known as neurotransmitters.The action potential causes some vesicles to fuse with the membrane of the presynaptic cell, thereby opening the vesicles and so releasing their neurotransmitters into the synaptic cleft.The neurotransmitters diffuse across the gap and bind to receptor sites on the neighboring postsynaptic neuron.This results in the opening of nearby ion channels in the postsynaptic cell membrane, causing ions to rush in or out and changing the local transmembrane potential of the cell and. either depolarization (an excitatory postsynaptic potential) or hyperpolarization (an inhibitory postsynaptic potential) occurs. A depolarization makes it more likely that a postsynaptic action potential will be generated. A hyperpolarization makes it less likely that a postsynaptic action potential will be generated.
Usually an excitatory action potential in the presynaptic neuron will trigger an action potential r in the postsynaptic cell.However, at a weak synapse, the excitatory postsynaptic potential will not produce an action potential. Some neurons form synapses with many others and receive synaptic inputs from many others. When action potentials fire simultaneously in several neurons that weakly synapse on a single cell, they may initiate an impulse in that cell even though the synapses are weak. This process is known as summation. On the other hand, a presynaptic neuron can release an inhibitory neurotransmitterdecreasing the postsynaptic neuron’s excitability and so decreasing its likelihood of firing an action potential. In this way, the output of a neuron may depend on the input of many others, each of which may have a different degree of influence, depending on the strength of its synapse with that neuron.Whether a synapse is excitatory or inhibitory is a function of the type of receptors and neurotransmitter at the synapse.
The action of neurotransmitters is stopped in four different ways. The neurotransmitter does not stay bound forever: Sooner or later it is shaken loose by random temperature-related vibrations. It can diffuse away from the synaptic cleft and so no longer act on the receptor. A specific enzyme can change the structure of the neurotransmitter so it is not recognized by the receptor. Glial cells, namely – astrocytes, can remove neurotransmitters from the synaptic cleft. Finally, they can be taken up by the presynaptic cell and then processed to be released for a later action potential. This is a common way that the action of norepinephrine, dopamine and serotonin is stopped. The time of these “clearing” processes ranges from a few tenths of a millisecond for the fastest, to several seconds for the slowest.
Another type of synapse, called an electrical synapse, is found throughout the nervous system, but is less common than chemical synapses.The electrical synapse is formed by a narrow gap between the pre- and postsynaptic neurons known as a gap junction. At a gap junction the cells are about 3.5 nm apart, rather than the 20 to 40 nm that separates cells at chemical synapses.The postsynaptic potential in electrical synapses is not caused by the opening of ion channels by chemical transmitters, but by direct electrical coupling between both neurons. Electrical synapses are therefore faster and more reliable than chemical synapses.They play an important role in reflex action actions –for example, withdrawing your hand quickly from a hot stove.The term synapse alone will be used to denote a chemical synapse.
Types of Neurotransmitters
Some of the properties that define a chemical as a neurotransmitter are difficult to test experimentally. It is easy using an electron microscope to recognize vesicles on the presynaptic side of a synapse. However, it may not be easy to determine the contained chemical or all of its properties. In order to determine if a chemical can be classified as a neurotransmitter, scientists, in the 1960s, proposed the following criteria:
There are precursors and/or synthesis enzymes located in the presynaptic neuron. The chemical is present in the presynaptic element. It is available in sufficient quantity in the presynaptic neuron to cause a biological effect in the postsynaptic neuron.When a neuron is stimulated (depolarized), a neuron must release the chemical. There are postsynaptic receptors and the chemical is able to bind to them. A biochemical mechanism for inactivation is present. If the chemical is applied on the post-synaptic membrane, it should have the same effect as when it is released by a neuron.
Not all of the neurotransmitters may actually meet every one of these criteria. Nitric oxide (NO) is not stored in synaptic vesicles. Rather, NO is released soon after it is produced and diffuses out of the neuron. NO then enters another cell where it activates enzymes for the production of “second messengers.”
Scientific advances have reduced the importance of these rules. Experiments that may have taken several years in the 1960s can now be done in a few months. It is unusual for the identification of a chemical as a neurotransmitter to remain controversial for very long.
Some examples of types of neurotransmitters appear in Table 2. The total number of neurotransmitters is not known, but is likely to be well over 100.
| Small Molecules | Amino Acids | Peptides | Gases |
| Acetylcholine (ACh) | Gamma-aminobutyric acid (GABA) | growth hormone-releasing hormone | Nitric Oxide (NO) |
| Serotonin (5-HT) | Aspartate | bradykinin | Carbon Monoxide |
| Dopamine (DA) | Glycine | calcitonin | |
| Histamine | Glutamate | gastrin | |
| Norepinephrine (NE) | secretin | ||
| Epinephrine | oxytocin | ||
| beta-endorphin | |||
| enkephalin | |||
| substance P | |||
| glucagon | |||
| insulin |
Table 2 Some Types of Neurotransmitters
3. The Nervous System
The nervous system consists of the central nervous system (CNS), the brain and spinal cord, and the peripheral nervous system (PNS). The PNS is the collection of nervous structures (nerves and ganglions) that do not lie in the CNS. A ganglion is a mass of nerve tissue containing the cell bodies of neurons.
The PNS has two divisions. The motor (efferent) division consists of motor nerves that carry impulses from the CNS to effectors (muscles and glands). The sensory (afferent) division consists of somatic and visceral sensory nerves that conduct impulses from receptors to the CNS.
Motor nerves, depending on their function, belong to two distinct systems: the somatic nervous system consisting of somatic (voluntary) nerves that conduct impulses from the CNS to skeletal muscles and the autonomic nervous system (ANS) comprised of visceral motor (involuntary) nerves that carry impulses from the CNS to smooth muscles, cardiac muscles, and glands.
The ANS regulates the activity of smooth and cardiac muscle and glands. It is composed of the sympathetic and parasympathetic nervous systems. The two subdivisions serve the same organs with different effects. The sympathetic division is the “fight-or-flight” subdivision, which prepares the body to cope with some threat. For example, if frightened, its activation results in increased heart rate and blood pressure. The parasympathetic division is the “rest and digest” system, functions with actions that do not require immediate reaction and is in control most of the time.
The nervous system and its divisions are summarized in Figure 2.
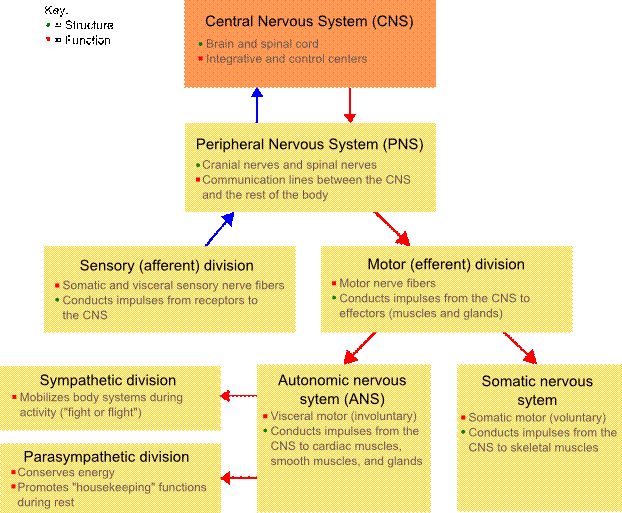
Figure 2 Divisions of the Nervous System
The activities of the two divisions of the ANS cannot always be ascribed to “fight” or “rest” situations. For example, standing up from a reclining or sitting position would result in dizziness or fainting due to a drop in blood pressure (i.e. – orthostatic hypotension), if not for a compensatory increase in the arterial sympathetic tonus. Another example is the constant modulation of heart rate by sympathetic and parasympathetic activity. More generally, these two systems should be seen as permanently modulating vital functions, in usually antagonistic fashion, to achieve homeostasis. Some typical actions of the sympathetic and parasympathetic systems are listed in Table 3.
| Sympathetic | Parasympathetic |
| Dilates pupils | Constricts pupils |
| Decreases salvation | Increases salvation |
| Increases respiratory rate | Decreases respiratory rate |
| Increases heart rate | Decreases heart rate |
| Constricts blood vessels | Dilates blood vessels |
| Inhibits digestion | Stimulates digestion |
| Relaxes bladder muscles | Contracts bladder muscles |
| Inhibits defecation | Stimulates defecation |
Table 3 Some Actions of the Sympathetic and Parasympathetic Nervous Systems
3. Autonomic Nervous System
Both the parasympathetic and sympathetic systems contain myelinated preganglionic nerve fibers that usually synapse with unmyelinated postganglionic fibers.Sympathetic Nervous System
The preganglionic motor neurons of the sympathetic system arise in the spinal cord. They pass into sympathetic ganglia forming two chains that run parallel to and on either side of the spinal cord. The preganglionic neuron may do one of the following in the sympathetic ganglion:
(a) Synapse with postganglionic neurons which then reenter the spinal nerve and so go to the sweat glands and the walls of blood vessels.
(b) Pass up or down the sympathetic chain and finally synapse with postganglionic neurons in a higher or lower ganglion. Then, it leaves the ganglion traveling to special ganglia (e.g. the solar plexus) in
the viscera. Here, it may synapse with postganglionic sympathetic neurons in the smooth muscular walls of the viscera.
(c) The preganglionic neuron in (b) may pass right through this second ganglion and into the adrenal medulla. where it synapses with the highly-modified postganglionic cells that make up the secretory portion of the adrenal medulla.Parasympathetic Nervous System
The preganglionic fibers of the parasympathetic system derive from the neural cell bodies of the motor nuclei of the occulomotor (cranial nerve: III), facial (VII), glossopharyngeal (IX), and vagal (X) cranial
nerves. There are also contributions from cells in the sacral segments of the spinal cord. These cranio-sacral fibers generally travel to a ganglion that is located near or within the target tissue. Because of
the proximity of the ganglia to the target tissue or organ, the postganglionic fibers are much shorter. For example, enteric system ganglia, innervate the digestive tube and are located inside its walls. It contains as many neurons as the entire spinal cord, including local sensory neurons, motor neurons and interneurons. It is the only truly autonomous part of the ANS. The digestive system can function well even in isolation. For that reason the enteric nervous system has been called “the second brain”.
The neurotransmitters released by the preganglionic and postganglionic sympathetic neurons are acetylcholine (Ach) and noradrenaline (or norepinephrine), respectively. The action of noradrenaline on a particular gland or muscle is excitatory is some cases, inhibitory in others (e.g. – inhibits peristalsis in the gastrointestinal (GI) tract).
Activation of the sympathetic system can produce many different effects because a single preganglionic neuron can synapses with many postganglionic neurons. Moreover, the release of adrenaline from the
adrenal medulla into the blood ensures that all the cells of the body will be exposed to sympathetic stimulation even if no postganglionic neurons reach them directly.
The chemical most commonly found in both pre and postganglionic synapses in the parasympathetic system is the neurotransmitter acetylcholine
Parasympathetic stimulation of the pupil from fibers derived from the occulomotor (Cranial nerve: III), facial (VII), glossopharyngeal (IX) nerves constricts or narrows the pupil. This reflexive action is an
important safeguard against bright light that could otherwise damage the retina. Parasympathetic stimulation also results in increased lacrimal gland secretions (tears) that protect, moisten, and clean the
eye. The vagus nerve (cranial nerve X) carries fibers to the heart, lungs, stomach, upper intestine, and ureter. Fibers derived from the sacrum innervate reproductive organs, portions of the colon, bladder, and rectum.
4. Somatic Nervous System
The pyramidal motor system controls all voluntary movements, through several nerve tracts. The corticospinal tract, runs from the cortex to the spinal cord and controls all the voluntary movements of the body. It is a two neuron system consisting of upper motor neurons in the primary motor cortex and lower motor neurons in the anterior horn of the spinal cord. 80 – 90 % of corticospinal tract axons cross to the other side in the distal medulla, just above the spinal cord. This is known as the pyramidal decussation. The corticospinal fibers in this new location, are now called the lateral corticospinal tract. The fibers which did not cross at the decussation form the anterior corticospinal tract. The upper motor neuron axons then synapse on lower motor neurons in the anterior horn of the spinal cord. The axons of these lower motor neurons then exit the spinal cord via the ventral root. The ventral root then joins the dorsal root to form the spinal nerve, which innervates the skeletal muscle. Fine finger movements and tap dancing are performed via the lateral corticospinal tract. Gross movements, like push-ups rely more on the anterior corticospinal tracts.
Muscles of the face and head are controlled by upper motor neurons located near the lateral fissure of the brain. These axons coalesce to form the corticobulbar tract. The upper motor neuron axons then synapse on Lower motor neurons of the cranial nerve nuclei which are located in midbrain, pons and medulla.
The extrapyramidal system is a network of nerves involved in the coordination of movement. The adjective extrapyramidal is used to distinguish it from the tracts of the motor cortex that travel through the “pyramids” of the medulla. Extrapyramidal tracts are mainly found in the reticular formation of the pons and medulla, and target neurons in the spinal cord involved in reflexes, locomotion, complex movements, and postural control. Its nuclei include the substantia nigra, caudate, putamen, globus pallidus, thalamus, red nucleus and subthalamic nucleus. All of these nuclei are synaptically connected to one another in the brainstem, cerebellum and the pyramidal system. Many of these synaptic connections are extremely complex and are still not completely understood. Extrapyramidal neurodegenerative disorders, such as, Parkinson’s disease and hemiballism affecting the substantia nigra and, unilaterally, the subthalamic nucleus, respectively, have yielded knowledge about its normal function.
5. Sensory Division
Sensory Body Pathways
The discriminative touch system detects pressure, vibration, and texture. This system relies on four different receptors in the skin (Meissner’s corpuscles, Pacinian corpuscles, Merkel’s disks, and Ruffini endings. The primary afferents ascend all the way to the medulla, on the ispilateral side of the cord, in the posterior columns. The secondary afferents cross in the medulla and ascend as the medial lemniscus. In the thalamus they synapse in the VPL (the ventroposterior lateral nucleus) and finally ascend to the sensory cortex. Itch and crude touch ascend by the paleospinothalamic pathway.
The pain and temperature system uses nociceptors in skin, muscle, bone, and connective tissue to detect temperature and pain. Nociceptors are free (bare) nerve endings that transduce a variety of noxious stimuli into an action potential, which is transmitted to their cell bodies in the dorsal root and trigeminal ganglia for the face. No nociceptors are found inside the CNS. Although pain will result from damage to a free nerve ending, in reality most pain is a result of substances released by damaged tissues: such as,prostaglandins, histamine, and substance P, which affect the nociceptors.
Sensations of pain an temperature are carried by the spinothalamic pathway which is formed in the folloing manner. The noniceptors’ axons lead from the skin into the dorsal spinal cord ganglion where they synapse with secondary neurons, called tract cells, in the nucleus proprius. The primary axons may travel one or two segments up or down the cord before synapsing. Lissauer’s tract is the tract carrying these migrating axons. The axons of the tract cells cross over (decussate) to the other side of the spinal cord via the anterior white commisure and to the anterolateral corner of the spinal cord. As they travel up the brainstem, the tract moves dorsally. The neurons ultimately synapse with third-order neurons in several nuclei of the thalamus – including the medial dorsal, ventral posterior lateral, and ventral medial posterior nuclei. From there, signals go to the cingulate cortex, the primary sensory cortex, and insular cortex respectively.
Cortical Areas for Sensation of Pain
The multiple pathways involve multiple thalamic nuclei that project to multiple cortical areas. In addition to the sensory cortex, painful stimuli activate neurons in the rostral cingulate gyrus and the insula. Consequently, all pain sensation is not lost when the primary sensory cortex is damaged. Lesions of the primary sensory cortex can affect the quality of pain sensations and the ability to localize the exact location of the painful stimulus.
Pain Control
Opium and related compounds, such as morphine block pain by binding to opiate receptors. Scientists reasoned that if there was such a receptor in the body, perhaps the body used its own endogenous form of opium to control pain. This hypothetical compound was named “endorphin”, from endogenous-morphine. Later, an entire class of endogenous peptide neurotransmitters (endorphins, enkephalins, and dynorphins) was discovered that interacted with the opiate receptors
There are opiate receptors throughout the central nervous system. In the dorsal horn, they are located on the terminals of the primary afferents, as well as on the cell bodies of the secondary afferents. Opiate interneurons in the spinal cord can be activated by descending projections from the brainstem (especially the raphe nuclei and periaqueductal grey), and can block pain transmission at two sites. They can prevent the primary afferent from passing on its signal by blocking neurotransmitter release, and they can inhibit the secondary afferent so it does not send the signal up the spinothalamic tract.
The proprioceptive system provides the sense that indicates whether the body is moving with required effort, as well as where the various parts of the body are located in relation to each other. The proprioceptive system arises primarily from afferents from muscle spindles, Golgi tendon organs, and joint receptors entering the spinal cord. The axons travel for a short distance in the discriminative touch system, in the posterior columns. Within a few segments, the proprioceptive information moves out of the dorsal white matter and synapses. After synapsing it ascends without crossing to the cerebellum. Conscious proprioception is communicated by the posterior column-medial leminiscus pathway to the cerebrum and unconscious proprioception is communicated primarily via the dorsal spinocerebellar tract to the cerebellum.
There is actually a fair amount of mixing that goes on between the tracts. Some light touch information travels in the spinothalamic tract, so that lesioning the dorsal columns will not completely knock out touch and pressure sensation. The pain and temperature system, although it does ascend to somatosensory cortex, also has multiple targets in the brainstem and other areas.
Sensory Facial Pathways
The three modalities of sensation from the body: discriminative touch, proprioception, and pain and temperature are also present in the face. However, sensory input from the face does not enter the spinal cord, but enters the brainstem via the trigeminal nerve. Just as in the spinal cord, these three modalities have different receptors, travel along different tracts, and have different targets in the brainstem. Once the pathways synapse in the brainstem, they join the pathways from the body on their way up to the thalamus.
The Trigeminal Nuclear Complex
The cells in the brainstem forming the trigeminal nucleus is huge and stretch from midbrain to medulla. Most of the sensory fibers enter the trigeminal ganglion, regardless of which trigeminal division they are coming from. Their cell bodies, like those of all the body sensory neurons, lie outside the CNS in the ganglion, and their proximal processes enter the brainstem in the mid-pons. From there they diverge to eir different targets. Each modality is described below.
Facial Discriminative Touch
The large diameter fibers enter directly into the main sensory nucleus of the trigeminal ( cranial nerve V), also called the principal nucleus. Just like the somatosensory neurons of the body; they synapse, and then cross. The secondary afferents can join the medial lemniscus on its way to the thalamus.
Facial Pain and Temperature
The small diameter fibers carrying pain and temperature enter at mid pons, and then do something unusual – they turn down the brainstem. They travel down the pons and medulla until they reach the caudal medulla, which is where they finally synapse and cross.
The tract that the descending axons travel in is called the spinal tract of V, and the long tail of a nucleus that they finally synapse in is called the spinal nucleus of V. These names come from the fact that they actually reach as far down as the upper cervical spinal cord. The secondary afferents from spinal nucleus of V cross to the opposite side, and join the spinothalamic tract on its way to the thalamus.
Facial Proprioception
The proprioceptive axons in the trigeminal nerve are the stretch and tendon receptors from the muscles of mastication, controlled by the facial nerve. These axons coming from the face have a strange characteristic unique among primary sensory neurons: their cell bodies are inside the CNS. though their cells look similar to cells in the dorsal root ganglion (the cell body does not come between the distal and proximal axon processes), they are located inside the brainstem in a nucleus called the mesencephalic nucleus. The mesencephalic nucleus is essentially a dorsal root ganglion that has been pushed into the CNS, so there are no synapses within it. The fibers enter the brainstem via a small branch of the trigeminal that bypasses the trigeminal ganglion, turn up towards the mesencephalic nucleus, pass by the cell body, and leave the nucleus immediately. Most then synapse in the nearby motor nucleus where they can initiate the stretch reflexes for the muscles of mastication.
Note on Facial Motor Control
Motor or efferent control is not considered a sensory modality, but it is the fourth component of the
extensive trigeminal complex. The motor nucleus of V lies just medial to the main sensory nucleus, and in it reside the a-motor neurons that control the muscles of mastication. The two principal muscles involved are the masseter (in your cheek) and the temporalis (over your temple), both of which tighten when you clench your teeth. The motor axons leave the mid-pons and bypass the trigeminal ganglion, and reach their targets via the mandibular division of the trigeminal nerve.
Pathways from the Thalamus
The somatosensory information from the face joins that from the body and enters the thalamus with it. However, face information actually enters a different nucleus in the thalamus. Recall that information from the body enters the ventroposterior lateral nucleus (VPL). Information from the face actually enters the ventroposterior medial nucleus(VPM). The thalamocortical afferents take all of the signals, whether from VPL or VPM, to primary somatosensory cortex. Once there, it is distributed in a somatotopic (body-mapped) fashion, with the legs represented medially, at the top of the head, and the face represented laterally.
Sensory System of the ANS
The sensory system of the ANS is composed of primary visceral sensory neurons found in three cranial sensory ganglia: the geniculate, petrosal and nodose ganglia, joined to cranial nerves VII, IX and X, respectively. These sensory neurons monitor the levels of carbon dioxide, oxygen and sugar in the blood, arterial pressure and the chemical composition of contents of the stomach and intestines and the sense of taste. The carotid body, innervated by the petrosal ganglion, is a small collection of chemosensors at the bifurcation of the carotid artery. It also senses the blood levels of oxygen and carbon dioxide.
Primary sensory neurons relay their impulses to secondary or relay sensory neurons through synapses. The first neuron releases a chemical neurotransmitter into a small space (the synapse) that is adjacent to second neuron, which influences the nerve impulse in the secondary neuron. These relay neurons are located in the medulla oblongata, forming the nucleus of the solitary tract (nTS), which integrates all visceral information. The nTS also receives input from a nearby chemosensory center, the area postrema, which detects toxins in the blood and the cerebrospinal fluid and is essential for chemically induced vomiting. It also ensures that an animal which has been sickened by a food never touches it again, called conditional taste aversion. The feedback from the sensory to the motor arm of visceral reflex pathways is provided by direct or indirect connections between the nucleus of the solitary tract and visceral motoneurons.
References
1. Guyton, A. C. Textbook of Medical Physiology, 6th edition, W. B. Saunders, 1980.
2. Neuroscience Tutorial, http://thalamus.wustl.edu/.
3. Neuroanatomy Web Resources. http://pathology.mc.duke.edu/neuropath/nawr/nawr_index.html
15 Responses to Scientific Qi Exploration-9a